

نورون دارای هسته، سیتوپلاسم و غشاء است. بخش هسته دار یاخته عصبی به نامهایپریکاریون perikaryon و سوما soma خوانده میشود. سیتوپلاسم سلول عصبی را نوروپلاسم میخوانند. در نوروپلاسم دانههای ریبوزوم ribosome ، میتوکندری mitochondria، دانههای لیزوزوم lysosome و دستگاه گلژی golgi، گویچههای چربی و مواد رنگی وجود دارد. میتوکندریها انرژی لازم را برای زندگی سلول فراهم میکنند .
دانههای ریبوزوم و شبکه آندوپلاسمی در پروتئین سازی و نقل و انتقال آنها نقش دارد و در برخی نواحی دانههای درشتی را میسازند که رنگهای قلیایی را به خود جذب میکنند و اجسام نیسل خوانده میشوند . دانههای لیزوزوم محتوی آنزیمهای هستند که میتوانند ذرات خارجی و مواد زاید را تجزیه و تخریب کنند. دستگاه گلژی در غشاسازی دستهبندی ترشحات نورون شرکت دارند. در نورونهای یک قطبی و دوقطبی تشخیص آکسون و دندریت از هم کار آسانی نیست. رشتههای بسیار نازک نوروفیبریل neurofibril و لولههای بسیار باریک نوروتوبول neurotubule در نوروپلاسم وجود دارد که نوعی اسکلت نوروپلاسم هستند و به حفظ شکل نورون و تارهای عصبی کمک میکند.
در هسته نورون، شیره هسته (نوکلئوپلاسم) nucleoplasm ، یک یا چند هستک (نوکلئول)nucleole و تودههای کروماتین وجود دارند. نورونها از لحاظ طرز خارج شدن تارهای عصبی از جسم سلولی شان به سه گروه یک قطبی (monopolar)،دوقطبی (bipolar) و چندقطبی (multipolar)تقسیم میشوند .
در نورونهای یک قطبی آکسون و دندریت از یک ناحیه جسم سلولی خارج میشود . نورونهای یک قطبی در عقدههای نخاعی انسان که گیرندههای حسهای پیکری هستند دیده میشود ، نورونهای بیمهرگان غالباً یک قطبی میباشد. در نورونهای دوقطبی آکسون از یک ناحیه و دندریتها از ناحیه دیگر جسم سلولی بیرون میآیند، گیرندههای بویایی در مخاط زرد بینی و برخی نورونهای شبکیه چشم از این نوع هستند. در نورونهای چندقطبی که تعداد آنها بسیار زیاد است آکسون از یک ناحیه و دندریت از چند ناحیه دیگر جسم سلولی خارج میشوند. نورونهای حرکتی شاخه پیشین نخاع و نورونهای پورکینجه (پورکنژ) purkinje مخچه و نورونهای هرمی شکل قشر مخ چندقطبی هستند. در برخی نواحی دستگاه عصبی مرکزی نورونهایی را که فاقد آکسون هستند شناسایی شدهاند، این نورونها تحریکات عصبی را فقط به نورونهای مجاور خود منتقل میکنند .
آکسونها، میلین دار و بدون میلین هستند. میلین یک ماده پروتئینی و چربی فسفر دار سفیدرنگ است که بعضی آکسونها را به صورت یک غلاف ناپیوسته پوشانده است.آکسوپلاسم axoplasm بخشی از نوروپلاسم سلول عصبی که به درون آکسون وارد میشود . گرههای رانویه nods of ranvier به فاصلههای بدون پوشش میلین در آکسون گفته میشود .
مخروط یا پیاز آکسون : ابتدای آکسون که در مجاورت جسم سلولی قرار دارد مخروط یا پیاز آکسون نام دارد و فاقد میلین است . انتهای آکسون هم فاقد میلین است . میلین به صورت مارپیچ در اطراف آکسون ساخته میشود . میلین در آکسونهای اعصاب پیرامونی توسط سلولهای به نام شوانschwann و در مراکز عصبی به وسیله یک نوع از ساخت همبند عصبی به نام اولیگو دندروگلیا oligodendroglia ساخته میشود . آکسونهای میلین دار در اعصاب مهرهداران و آکسونهای بدون میلین در اعصاب بی مهرگان دیده میشود.
در درون دندریتهای میتوکندری ، شبکه درون سیتوپلاسم، ریبوزمها و سایر ضمایم سیتوپلاسمی دیده می شود. سطح خارجی دندریتها دارای گیرنده های غشایی است که اطلاعات را از سایر نورنهای دیگر دریافت می کند. در درون دندریتهای میتوکندری برآمدگیهای ریزی وجود دارد که به نام خارهای دندریتیdendritic spines نامیده میشود . تارهای عصبی که پیامهای حسهای پیکری را از گیرندهها به نخاع میآورند تارهای طویلی هستند که اغلب غلاف میلین دارند و ظاهری شبیه آکسون دارند ،اما در واقع دندریت هستند.
غشا در حالت آرامش یعنی فقدان محرک خارجی قطبی (پولاریزه) Polarized است یعنی داخل غشاء بار منفی و بیرون غشاء بارمثبت دارد و اختلاف سطح الکتریکی بین درون و بیرون غشاء برابر است . پتانسیل غشا در سایر سلولها هم وجود دارد اما در سلولهای عصبی و عضلانی از سایر سلولها زیادتر است. میزان پتانسیل آرامش غشا در تارهای عصبی ۷۰ میلی ولت و در تارهای عضلانی ۹۰ میلی ولت است.
چگونگی ایجاد پتانسیل آرامش غشای عصبی
تعداد یونهای مثبت در سطح بیرونی غشا کمی بیشتر از یونهای منفی و تعداد یونهای منفی در درون غشا کمی بیشتر از یونهای مثبت باشد، در ایجاد این نابرابری یونها چندعامل شرکت دارند: ۱)پمپ سدیم -پتاسیم ۲) تفاوت نفوذپذیری غشاء در حالت آرامش نسبت به یونهای سدیم Na و پتاسیم k. در حالت آرامش، نفوذپذیری غشاء سلول عصبی نسبت به یونهای پتاسیم k در حدود ۱۰۰ برابر بیش از یونهای سدیم Na است. یونهای سدیم Na و کلرcl مهمترین یونهای محیط خارجی و یونهای پتاسیم k و پروتئینها و مولکولهای آلی فسفردار مهمترین مواد محیط داخلی تارهای عصبی هستند. ترکیبات چربی غشا به صورت یک عایق در بین بارهای مثبت و منفی قراردارد.
پمپ سدیم –پتاسیم
در غشای سیتوپلاسمی همه سلولهای بدن، پدیده انتقال فعال یونهای سدیم و پتاسیم وجود دارد که پمپ سدیم- پتاسیم نامیده می شود. مجموعه ای از آنزیمهای پروتئینی با صرف انرژی، یونهای سدیم Na+ را به بیرون و یونهای پتاسیم K+را از بیرون به داخل سلول منتقل می کنند. سه یون سدیم Na+ با دو یون پتاسیم K+معاوضه می شوند. یون هایی سدیم Na+ و پتاسیم K+هر دوبار مثبت دارند، اما با توجه به اینکه سه یون مثبت Na+ با دو یون مثبت K+معاوضه می شود، پمپ سدیم – پتاسیم تعادل بین تعداد یونها مثبت و منفی را در بیرون و درون نورون بر هم می زند و باعث افزایش یونهای مثبت خارج Na+ نسبت به یونهای مثبت داخل K+ می شود، به این خاطر پمپ سدیم ، پتاسیم را پمپ الکتروژنیک electrogenic pump می نامند. پتانسیل الکتریکی غشاء که مستقیماً براثر پمپ سدیم – پتاسیم ایجاد می شود فقط چندولت است، و بخش زیادی از پتانسیل آرامش غشاء نورون نتیجه انتشار یونهای پتاسیم به خارج است. پمپ سدیم- پتاسیم باعث می شود تا همواره تراکم پتاسیم در درون تارعصبی بیش از محیط خارج آن (مایع میان بافتی بدن)و تراکم سدیم در محیط خارجی تارعصبی بیش از محیط درون آن باشد. غشای عصبی در حالت آرامش نسبت به انتشار و جابه جایی پتاسیم K+ نسبتاً نفوذپذیر و نسبت به سدیم Na+ تقریباً نفوذناپذیر است.
در حالت استراحت فقط خروج تعداد بسیار اندکی از یونها پتاسیم (یک یون از هر صدهزار یون داخلی) برای ایجاد پتانسیل آرامش کافی است، با خروج پتاسیم K+ یک نیروی الکتریکی در جهت جلوگیری از خروج پتاسیم بیشتر دارد عمل می شود و در این حالت بین نیرو، انتشار که پتاسیم را به خارج می راند و دافعه یونهای مثبت که مانع خروج یونهای مثبت پتاسیم K+می شود ، تعادل برقرارمی شود،از این روبه پتانسیل آرامش غشا ، پتانسیل تعادل هم می گویند. فقط غشای سلولهای عصبی و عضلانی می توانند پتانسیل عمل تولید کنند، به همین جهت غشاء این دو نوع سلول را تحریک پذیر می گویند.
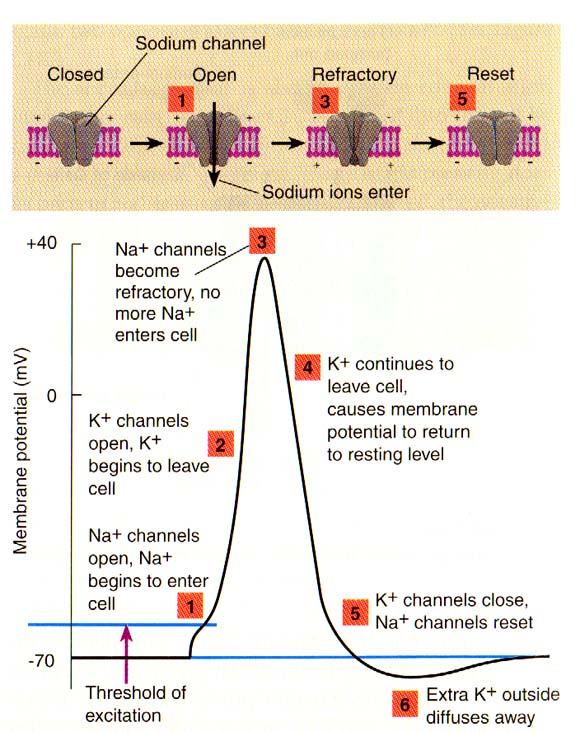
مراحل پتانسیل عمل Acting Potential
۱-پتانسیل آرامش :mv ۷۰ – Resting Potential
۲-پولاریزه mv ۵۵ – Polarization
۳-دپولاریزاسیون mv ۴۰ + تا ۵۵ – Depolerization
۴-رپولاریزاسیون mv ۷۰- تا ۴۰+ repolarization
۵-هیپر پولاریزاسیون Hyperpolarization
پتانسیل موضعی (محلی)
باز و بسته شدن دریچه های سدیم در غشاهای تحریک پذیر به ولتاژ غشا وابسته است، مواد بی حس کننده موضعی مانند نوکائین Novocain و گریلوکائین Xylocain به جلوگیری از ورود یونهای سدیم به تارعصبی مربوط است یعنی مانع دپولاریزاسیون غشا و در نهایت پتانسیل عمل می شوند. اما حشره کشها با ازکار انداختن پمپ سدیم باعث می شوند تا دریچه های سدیم باز بمانند و با وجود فقدان محرک، امواج عصبی درونزاد تولید شود و یونهای سدیم در درون تارعصبی تجمع یافته و غشاء عصبی به پتانسیل آرامش نرسد، کاهش کلسیم cl از دلایل آن است.
آستانه تحریک و قانون همه یا هیچ
کمترین شدت محرکی که بتواند غشای عصبی را تحریک کرده و پتانسیل عمل دران ایجاد کند، آستانه تحریک خوانده می شود. این حد از پتانسیل آرامش را سطح آتش یا سطح شلیک پتانسیل عمل می نامند. در یک عصب که خود از تعدادی تارعصبی ساخته شده است قانون همه یا هیچ صدق نمی کندزیرا با افزایش شدت محرک به تدریج تعداد بیشتری از تارهای عصبی تحریک می شوند و یا سطح کلی ثبت شده شدت بیشتری می یابد.
شدت پایه و زمان مفید
بین شدت محرک و مدت آن رابطه معکوس وجود دارد، در صورتی که زمان محرک از حدی بیشتر شود آستانه تحریک به حداقل خود می رسد و از آن پس ثابت می ماند. این مدت محرک را که در آن شدت آستانه به کمترین میزان خود می رسد زمان مفید می گویند.
مراحل تحریک ناپذیری Refrectory Period
مرحله اول ، تحریک ناپذیری مطلق ، یک تا دو هزارم ثانیه ، آستانه تحریک تار عصبی بی نهایت و تحریک پذیری آن صفر است که منجر به دپولاریزاسیون می شود. مرحله دوم، تحریک ناپذیری نسبی، ده تا پانزده هزارم ثانیه، تا پس از رپولاریزاسیون ادامه دارد.
از لحظه ای که محرک بر تار عصبی وارد می شود تا لحظه ای که پتانسیل عمل آغاز می شود، زمان نهفته lacent Period گفته می شود. پتانسیل عملی که در محل تحریک بوجود می آید با شدت یکسان و بدون کاهش در طول تار عصبی سیر می کند. هر منطقه تحریک شده باعث تحریک ناحیه مجاور شده ،خود به حالت آرامش می رسد. موج عصبی در هدایت خود به نواحی دور دست کاهش نمی یابد و شدت آن در ابتدا و انتهای تارعصبی یکسان است. در بدن با توجه به اینکه در حالت عادی پتانسیل عمل فقط در یک سر تار عصبی بوجود می آید ، سیر آن یک طرفی است .
سرعت هدایت موج عصبی در طول یک تار ثابت، ولی در تارهای مختلف متفاوت است. هر چه قطر تار عصبی بیشتر باشد، سرعت هدایت موج عصبی در آن زیادتر است. وجود میلین نیز باعث افزایش سرعت انتشار موج عصبی می شود. سرعت هدایت پتانسیل عمل در قطورترین تارهای میلین دار انسان در حدود ۱۲۰ متر در ثانیه و در نازک ترین تارهای بدون میلین تقریباً ۵/ متر در ثانیه است.
تارهای عصبی را با توجه به قطر آنها و وجود یا فقدان میلین در آنها به سه دسته تقسیم می کنند : گروه A (دارای میلین): قطورترین تارهای عصبی هستند و تارهای گروه آلفا قطورتر هستند ،فاصله گره های رانویه در آنها زیادتر است و سرعت سیرموج عصبی در این تارها بیشترین مقدار را دارد و زمان نهفته و مراحل تحریک ناپذیری آنها کوتاه است. گروه B(دارای میلین) سرعت جریان عصبی ۳ تا۱۵ متر در ثانیه و گروه C (فاقد میلین) سرعت جریان عصبی ۵/ تا ۲ متر در ثانیه است.
هدایت جهشی Saltatroy Conduction
وجود غلاف میلین در تارهای عصبی که به صورت یک عایق، غشای آکسون را می پوشاند باعث می شود تا فقط در گره های رانویه، پتانسیل آرامش تولید شود و در نتیجه فقط این گره ها قادر به ایجاد پتانسیل عمل باشند. به همین دلیل هدایت موج عصبی در تارهای عصبی میلین دار به صورت جهشی صورت می گیرد ، یعنی از یک گره به گره بعدی منتقل می شود، این نوع انتشار باعث افزایش سرعت موج عصبی وصرفه جویی در مصرف انرژی می شود، وجود مایع میان بافتی در سطح خارجی تارهای عصبی که هادی جریان الکتریکی است مدارخارجی انتقال تحریک از یک گره به گره بعدی را امکان پذیر می کند. بخش داخلی دیوار تحریک این گره ها به وسیله آکسوپلاسم در دل تار تأمین می شود. در تارهای عصبی میلین دار فقط گره های رانویه می توانند یونها را به میزان لازم برای ایجاد پدیده های الکتریکی از خود عبور دهندو میزان عبور یونها به علت کوچک بودن مساحت گره های رانویه بسیار کمتر از تارهای عصبی بدون میلین است.
پتانسیل عمل مرکب
هر عصب یا تنه ی عصبی از تعدادی تارعصبی ساخته شده است. با قراردادن الکترون های خارجی بر روی عصب و اتصال آنها به اسیلوسکوپ و تحریک عصب با محرک نسبتاً قوی می توان پتانسیل عمل مجموعه تارهای تحریک شده را ثبت کرد. با افزایش شدت محرک که به تدریج تحریک تارهای عصبی بیشتر می شود بر ولتاژ پتانسیل عمل مرکب نیز تا حد معینی افزوده می شود. در صورتی که محل ثبت پتانسیل عمل از محل تحریک فاصله کافی داشته باشد شکل پتانسیل عمل مرکب دو یا چند قله ای پیدا می کند که نشانه وجود تارهای عصبی مختلف با سرعت هدایت متفاوت در عصب است.
منبع : فیزیولوژی اعصاب و غدد درون ریز / سیدعلی حائری روحانی / نشر سمت / فصل ۳
🌿 اگر ذهنتان درگیر است، تنها نیستید
بسیاری از افراد در بخشهایی از زندگی خود با اضطراب، فشارهای روانی یا احساس سردرگمی روبرو میشوند. در چنین شرایطی، گفتوگو با یک درمانگر، میتواند کمک کند تا ذهن آرامتر و مسیر زندگی روشنتر شود. اگر احساس میکنید به همراهی و حمایت نیاز دارید، میتوانید برای مشاوره حضوری یا آنلاین پیام دهید.
📱 ارتباط با ما : ۰۹۳۵۵۷۵۸۳۵۸